
- Šta je uopšte evolucija
- Evolucija danas: mutacije i nastanak informacija
- Evolucija danas: selekcija i širenje informacija kroz populaciju
- Evolucija danas: prirodna selekcija, geni i populacije
- Teorija evolucije života na Zemlji
- Dokazi: Evoluciona istorija i ugnježdena hijerarhija
- Dokazi: Genetika, genomika i tragovi evolucije
- Dokazi: Fosilni zapis i razvoj života
- Dokazi: Razvojni i fiziološki tragovi
- Zakoni prirode ili slučajna konfluenca dokaza?
U prethodnom poglavlju, videli smo šta je to selektivni pritisak. Pogledajmo sada par primera efekata koje kombinacija mutacija i selekcije ima u prirodi.
Primer: leptiri, bakterije i brzina evolucije. Bakterije iz genusa Wolbachia parazitiraju na insektima, i po broju i proširenosti spadaju među najuspešne organizme na planeti Zemlji. Nekoliko vrsta ovih bakterija živi u leptirima, gde su razvili jedan veoma efektivan način da ubrzaju svoj prenos.
Naime, jaja koja legu zaražene ženke leptira takođe nose u sebi infekciju. Spermatozoidi mužjaka, međutim, ne prenose zarazu, i bakterije koje zaraze mužjake otud nikada ne uspevaju da ostave potomstvo: kada mužjak leptira umre, bakterije umiru zajedno sa njim.
Otud, ove vrste bakterija su evoluirale jedan otrovni protein: u reproduktivnim organima zaražene ženke, bakterije ovim proteinom selektivno ubijaju jaja iz kojih bi se inače izlegli mužjaci, ostavljajući jaja sa ženkama netaknutim. Ovaj proces dovodi do situacije u kojoj veoma malo mužjaka doživi da se ikada izlegne iz jajeta, i populacija leptirova se sastoji skoro u potpunosti od ženki (koje su skoro sve zaražene Wolbachiom).
Vrsta rasprostranjenog tropskog leptira Hypolimnas bolina je bila lep primer ovoga. Na Samoanskom ostrvu Savai, ženke ovog leptira su bile česte, dok su mužjaci činili manje od 1% ukupne populacije. No, 2005-te godine se desilo nešto novo: leptir je evoluirao novi gen, supresor, koji je onemogućio delovanje bakterijskog toksina. U roku od samo godinu dana, mužjaci ove vrste su odjednom postali toliko česti da su činili 39% ukupne populacije na ostrvu.

Ovo je, naravno, očigledan rezultat procesa o kojima smo govorili u prethodnim poglavljima. Mutacije su proizvele novu informaciju, nova informacija je potpala pod snažan selektivni pritisak koga na leptire vrše bakterije, i otud se odjednom proširila kroz populaciju.
Ovo nam ukazuje na jedan važan princip: novi selektivni pritisci veoma brzo menjaju genetski sastav populacije. Ovo je tačka koju vredi dodatno razjasniti.
Uzmimo radi toga još jedan dokumentovan primer: miševe (vrsta Chaetodipus intermedius) koji su se naselili na području koje je većim delom prekriveno vulkanskim pepelom. Miševi sa bledom bojom krzna su jasno vidljivi na tamnoj pozadini, i predstavljaju lak plen za grabljivce. Ovo proizvodi snažan pritisak za tamnije krzno.
I zaista, ovi miševi su brzo razvili sve tamnije i tamnije krzno, dok na kraju nisu postali crni, boje koja je veoma slična boji peska u kome žive. Ovo je sada granica adaptacije: ako se krzno uklapa sa peskom, više ne može da evoluira da se “još bolje” uklapa sa peskom (ne postoji neka još bolja, “crno-smeđija” boja). Mutacije će u budućnosti i dalje proizvoditi varijacije u boji krzna, ali sve boje osim ove ustanovljene će biti eliminisane prirodnom selekcijom; samim tim, boja krzna se neće dalje menjati.
Ovo nas dovodi do razloga za relativnu stabilnost mnogih osobina vrsta. Kada se vrsta jednom adaptira na promenu u okolini, ta adaptacija ostaje stabilna, sve dok neka nova promena ne proizvede novi selektivni pritisak.
Otud, organizmi koji žive u stabilnim uslovima (tzv. stabilnim ekološkim nišama) će se menjati veoma sporo (primer ovoga su dubokookeanske krabe, čija se životna okolina nije promenila stotinama miliona godina; današnje krabe su veoma slične, mada ne identične, vrstama koje su postojale pre više stotina miliona godina). Sa druge strane, organizmi koji žive u promenjivim sredinama će se menjati mnogo brže.
Možemo sada ovde da postavimo jedno veoma važno pitanje: koje su verovatnoće ovde u pitanju? Genomi su veliki, i šansa da mutacija pogodi baš određeno mesto je naizgled veoma mala. Ovo je kod mnogih ljudi veliki kamen spoticanja, zbog čega vredi ilustrovati vremenski okvir i veličinu populacije nad kojom ovi procesi deluju. Pošto su današnji primeri veoma matematički kompleksni, uzmimo jednu uprošćenu rekonstruisanu verziju.

Teorijski primer: mimikrija i krila leptira. Mnogim pticama su insekti glavni izvor hrane, zbog čega kod insekata postoji snažan selektivni pritisak da razviju načine kojima mogu da se odbrane od ptica. Neki su razvili otrovne ili neukusne supstance u telu, drugi su se adaptirali tako da se uklapaju što bolje mogu u svoju okolinu… Ali postoje i neka veoma zanimljiva rešenja. Recimo, nekoliko vrsta leptira je razvilo mrlje na krilima koje veoma liče na oči sove. Male ptice koje se hrane leptirima se veoma boje sova, i otud umesto da pojedu leptira, one se uplaše i pobegnu. Na gornjoj slici se može videti jedan ovakav leptir, zvani Caligo memnon.
No, kako ovako nešto može da nastane? Šanse da slučajno nastanu mrlje koje liče na oči sove su veoma male, reda jedan u deset milijardi! Svakako, tako nešto se ne može zaista desiti, zar ne?
U stvari, itekako može, ako pravilno razumemo količinu vremena i broj jedinki koje su uključene u proces evolucije. Uzmimo neke prosečne cifre u razmatranje.
Uobičajena populacija neke vrste leptira u određenoj sredini sadrži oko sto miliona jedinki. Prosečan leptir ostavi oko dvadesetak potomaka, što nam daje dve milijarde potomaka po generaciji. Uzmimo u obzir da svaki novi leptir nosi po, da zaokružimo, sto novih mutacija. Ovo je dve stotine milijardi genetskih promena u svakoj generaciji.
Ako u ovo ubacimo nekoliko tehničkih detalja – veličinu genoma prosečnog leptira (oko sto miliona genetskih “slova”, tj. tačnije DNK baznih parova), i broj gena koji kontrolišu izgled krila (oko pet ili šest) – možemo da očekujemo da će se u svakoj generaciji roditi sto četrdeset hiljada jedinki koje imaju promenjen izgled mrlja na krilima!
Mi ovo retko primećujemo, pošto je sto četrdeset hiljada od dve milijarde novorođenih leptirića samo 0,007% cele populacije; dakle, praktično svaki leptir na koga naletimo će imati “normalan” izgled krila, i samo jedan u svakih petnaest hiljada će imati malčice promenjene mrlje. Ako povremeno i naletimo na nekog takvog, pomislićemo da je u pitanju prosto neobičan primerak (što u stvari i jeste tačno).
Razmislimo sada šta ovo znači u kontekstu dužih vremenskih perioda. Sto četrdeset hiljada leptira po generaciji, jedna generacija godišnje. Za hiljadu godina, ovo je sto četrdeset miliona novih oblika krila. Za milion godina, ovo je sto četrdeset milijardi novih oblika mrlja na krilima.
Apsolutno najveći broj – 99,999% – ovakvih promena neće imati nikakvog efekta. Novi geni će se utopiti u “pozadinu” dominantnih gena u okviru populacije, i pre ili kasnije će nestati. No, ovaj nestanak nije trenutan. Mada će u svakoj generaciji od onih 140.000 većina uginuti bez potomstva, i dalje će ostati desetine hiljada koji će ostaviti potomstvo i tom potomstvu ostaviti svoje osobine. Time će uvek postojati određen deo vrste – možda 0,01% ukupno – koji ima značajno drugačije mrlje na krilima nego što je uobičajeno.
I sad, jednog dana jedan takav leptir ima mrlje koje u određenom svetlu pod određenim uglom liče na lice sove. Ovo leptiru daje veoma slabu zaštitu – povremeno, možda jednom u dvadeset puta kada potomka takvog leptira napadne ptica, ta ptica se uplaši. Potomci ovog leptira će samo povremeno zvog ovih mrlja preživeti u situaciji u kojoj inače ne bi preživeli. Ali ovo znači selektivnu prednost! Ova osobina će se sada neizostavno širiti kroz populaciju. Iz generacije u generaciju, sve će više leptirova nositi ove nove osobine – hiljade, desetine hiljada, stotine hiljada…
A kao što smo videli na prethodnim primerima, povećanje broja jedinki koje nose određenu osobinu znači da postoji povećana šansa da će se ta osobina dalje razvijati. Dok se ova osobina širi kroz populaciju, povremeno će potomci leptira sa novim mrljama imati nove mutacije koje dodatno menjaju mrlje. Ako se mrlje izmene tako da sada manje liče na sovu, ovi potomci gube selektivnu prednost – a gubitak selektivne prednosti je isto što i negativna mutacija, i postaje predmet pritiska koji te gene eliminiše iz populacije.
Ali ako nova dodatna promena proizvede mrlje koje još više liče na sovu, i još bolje plaše ptice…ta nova promena sada dobija još veću selektivnu prednost i još brže se širi kroz populaciju.
U roku od par stotina hiljada generacija, mrlje dostižu granicu adaptacije: postaju onoliko slične sovi koliko je to moguće na krilima ove vrste leptira. Procesima mutacije i selekcije nad velikim brojem jedinki preko dugog perioda vremena, proizveli smo nešto što na prvi pogled izgleda neverovatno ili nemoguće.
Dakle, selektivni pritisak upravlja razvojem novih osobina, određuje brzinu njihovog razvoja, i može vremenom da proizvede naizgled neverovatne stvari. No, šta se događa kada postojeći selektivni pritisak bude uklonjen? Pogledajmo još jedan primer.

Primer: oči pećinskih riba. Na slici iznad možete da vidite jednu vrstu pećinske ribe, Amblyopsis spelaea. Odmah se može videti je jedan veliki nedostatak: ova riba (kao i mnogi srodni organizmi) uopšte nema oči.
U okviru populacije rečnih riba, vid je veoma važna osobina. Ako se među potomcima neke ribe rode pojedinci sa mutacijama koje izazivaju slepilo (što se u stvari događa relativno često, jedna ribica u svakih par hiljada se rađa slepa), taj potomak neće dugo preživeti: biće lak plen za grabljivce, i mnogo teže će nalaziti hranu nego njegova sabraća. Otud, na ribe u rečnom ekosistemu deluje selektivni pritisak za održanje vida. Pošto mutacija koja uništava vid ide protiv ovog selektivnog pritiska, ona je snažno negativna.
U pećinama, međutim, situacija je potpuno drugačija. Pošto nema svetlosti, nema ni selektivnog pritiska za održanje vida: jedinke koje vide nemaju nikakvu prednost nad slepim jedinkama.
Oči kičmenjaka troše energiju neprestano, čak i u potpunom mraku; otud, za ribu u pećini, oči predstavljaju neprestano ulaganje energije bez ikakvog dobitka. Ako se potomak ribe rodi sa mutacijama koje dovode do slepila, on u ovom slučaju ima prednost: ne ulaže neprestano energiju u nepotreban organ. Pritom, pošto nema selektivnog pritska (jedinke sa vidom nemaju nikakvu prednost), on ništa nije izgubio
Ovo je jedan od mnogih primera važnosti selektivnog pritiska za održavanje postojećih osobina. Ako se ukloni selekcija, ne samo što nema nastanka i širenja informacija, već postojeće informacije neizbežno degenerišu.
Dakle, danas možemo da posmatramo kako procesi mutacija unose varijacije, i kako procesi selekcije menjaju genetski sastav organizama.
Pošto genetski sastav definiše vrstu, njegova promena znači automatski i promenu vrste: ako dovoljno promenimo gene, promenićemo i vrstu. U prirodi možemo uživo da primetimo različite nivoe ovakvih promena. Recimo, konji i magarci su potomci iste vrste koja je podeljena na dva dela; ta dva dela su zatim bila podložena različitim selektivnim pritiscima. Rezultat nakupljenih genetskih razlika je da ove dve vrste više nisu u stanju da proizvedu plodno potomstvo. Ukrštanje konja i magarca proizvodi sterilne potomke (mazge ili mule) koji nisu u stanju da se dalje razmnožavaju. Ovaj proces takođe možemo uživo da gledamo na nekoliko primera u prirodi, među kojima su možda najbolji takozvane prstenaste vrste.
Primer: vrste i sojevi zelenog slavuja. Zeleni slavuj (Phylloscopus trochiloides) je ptica koja živi u planinskim šumama, i potiče iz oblasti Himalaja (mesto označeno sa A na mapi ispod). Šireći se iz svoje početne oblasti, prelazeći sa planine na planinu, populacija zelenog slavuja se raširila u prstenu oko Tibetanske stepe (u kojoj nema dovoljno drveća da bi slavuj uspeo da preživi).
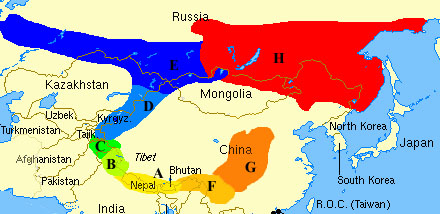
U toku ovog širenja, međutim, populacije slavuja u odvojenim područjima su postajale sve udaljenije jedna od druge. Ptice sa udaljenih planina su nakupljale mutacije, i populacije su postajale sve različitije i različitije. Boja i oblik perja su postali značajno različiti između odvojenih populacija.
Slavujeva pesma se polako menjala zajedno sa selidbom, što je dovelo do dodatne izolacije među populacijama: ženka jedne grupe više “ne razume” pesmu mužjaka druge grupe, i time je međusobno ukrštanje otežano.
Kakav je ukupan efekat ovih događaja? Susedne grupe su još uvek dovoljno genetski i vizuelno slične da bi mogle da se pare. Slavuji iz grupe A se redovno pare sa slavujima iz grupa F i B; ređe sa slavujima iz grupa C i G. Međutim, kada se udaljenost poveća, više nema parenja: parovi se nikada ne uspostavljaju između grupa E i A, ili F i D. A na samim ivicama prstena, dolazi do situacije u kojoj su genetske promene dovoljno velike da sprečavaju ukrštanje: slavuji iz grupa G i H ne mogu da se pare jedni sa drugima, jer takva paranja više ne mogu da proizvedu živo potomstvo. Genetske razlike su prosto suviše velike.
Posmatrajući zelenog slavuja, možemo videti mehanizam gradacije vrsta u akciji: susedne grupe još uvek pripadaju istoj vrsti, ali kada udaljenost postane suviše velika, moramo početi da govorimo o dve vrste slavuja.
Nastanak vrsta kroz reproduktivnu izolaciju je primećen i u laboratoriji. Jedan konkretan primer: 1964-te godine, na obali okeana u Kaliforniji, naučnici su pokupili pet-šest crva vrste Nereis acuminata. Od ove nekolicine crva, u laboratoriji su ovi crvi (radi raznih eksperimenata iz ćelijske biologije) razmnoženi u više hiljada jedinki. Četiri mužjaka i četiri ženke su ubrzo nakon toga poslati kolegama u laboratoriji na istoku Sjedinjenih Američkih Država, gde je njihovo potomstvo nastavilo da živi sledećih dvadeset godina. U toku tog perioda, ove dve populacije su bile izolovane jedna od druge.
Nakon dvadeset godina, dve populacije crva su nakupile toliko različitih mutacija da su postale reproduktivno nekompatibilne. Jedinke iz istočne grupe uopšte ne mogu da proizvedu potomstvo sa jedinkama iz zapadne grupe. Od jedne vrste, efektivno su nastale dve nove.
Ono što je važno je da nijedna pojedinačna mutacija ne proizvodi ovakve rezultate. Promene u vrstama su rezultat nakupljanja više različitih mutacija kroz mnogo generacija, pod različitim selektivnim pritiscima. Ovo nas dovodi do poslednje veoma važne stavke:
Jedinka doživljava mutacije, populacija doživljava selekciju.
U sledećem poglavlju, sabraćemo sve prethodno rečeno, i formulisaćemo pravila evolucije.
[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]