
Sadržaj:
- Uvod – šta su to prelazni fosili
- Razvoj višećelijskih organizama i Kambrijska eksplozija
- Razvoj kičmenjaka
- Od riba do vodozemaca
- Od vodozemaca do reptila
- Od reptila do sisara
- Od reptila do ptica
- Kratak pregled evolucije biljaka
- Žirafe, kitovi, konji i druge napomene
9. Žirafe, kitovi, konji i druge napomene
Za kraj ovog teksta, uložićemo malo vremena u obradu nekih čestih antinaučnih tvrdnji koje su povezane sa fosilnom istorijom života.
Vrat žirafe
Vrat žirafe je veoma upečatljiva pojava, za koju su prelazne forme su otkrivene tek u zadnjih desetak godina. Žirafe nikada nisu bile usamljena, odvojena grana, imaju bliske srodnike koji su opstali do danas: takozvane okapije. Ali mada vrat okapija sadrži mnoge adaptacije viđene kod drugih žirafida, on je mnogo kraći. Međustepeni su u međuvremenu otkriveni u vidu fosila Sivatherium, Bramatherium, i Samotherium, koji pokazuju postepen razvoj osobina koje vidimo kod modernih žirafida.
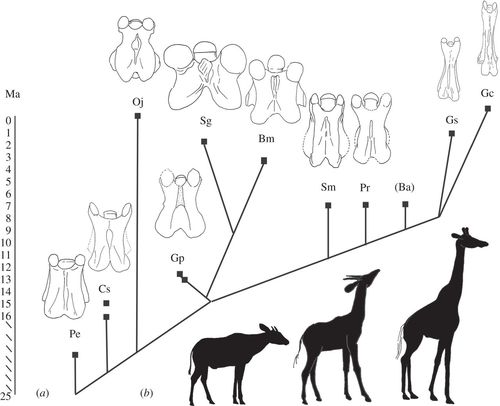
Kladogram koji pokazuje razvoj žirafida kroz vreme. Skala je u Ma (milionima godina pre sadašnjosti). Sheme pokazuju razvoj C3 vertebri sa leđne strane. Pe, Prodremotherium elongatum; Cs, Canthumeryx sirtensis; Oj, Okapia johnstoni; Gp, Giraffokeryx punjabiensis; Sg, Sivatherium giganteum; Bm, Bramatherium megacephalum; Sm, Samotherium major; Pr, Palaeotragus rouenii; Ba, Bohlinia attica; Gs, Giraffa sivalensis; Gc, Giraffa camelopardalis. Adaptirano iz Danowitz et al. R. Soc. open sci. 2150393.
Zanimljivo otkriće iz ovih fosila je da je produženje vertebri i glave počelo pre produženja vrata, što indikuje da je uzrok ove evolutivne adaptacije samo delimično bio ishrana. Mužjaci žirafa se bore za ženke tako što se međusobno udaraju vratom; moguće je da je seksualna selekcija bila najvažniji aspekt u razvoju velikih i snažnih vratova kod ovih vrsta? Niz novih fosila otkrivenih u toku zadnjih nekoliko godina, čija analiza je i dalje u toku (Schansitherium, Decennatherium) će nam dati još više podataka na osnovu kojih se ovo pitanje može rešiti. Štagod da je uzrok, međutim, jasno se u fosilima može videti postepen, korak po korak, razvoj modernih vratova žirafe. Samotherium stoji tačno na pola puta između vratova okapija i vrata modernih žirafa.
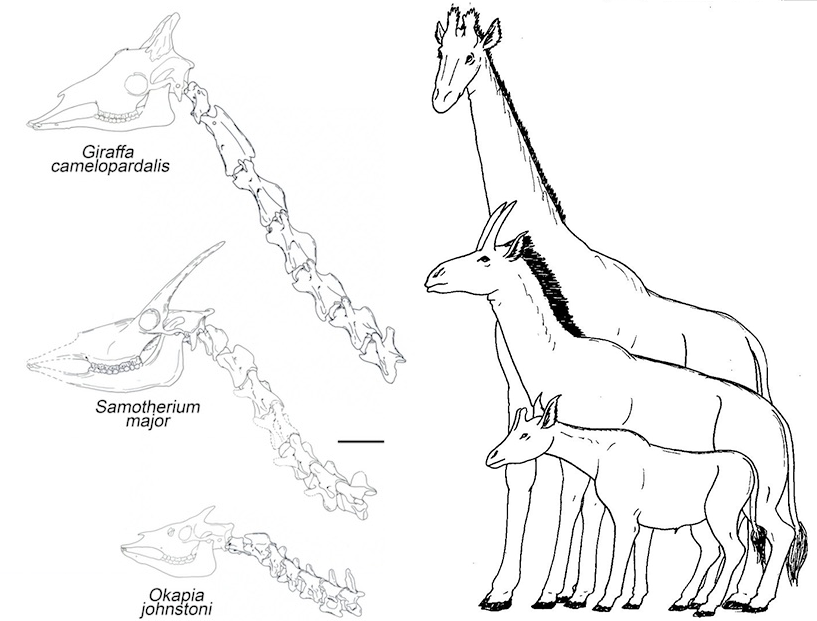
Levo: poređenje skeleta okapija, samoterijuma, i žirafe (Nikos Solounias).
Desno: izgled žirafe i okapija, i rekonstrukcija izgleda samoterijuma (autor: Apokryltaros).
Povrh ovih fosilnih dokaza, tu su i drugi jasni tragovi evolucije. Isto kao i svi drugi sisari, od miševa do kitova, žirafe takođe imaju sedam pršljenova u vratu. Ovi pršljenovi pokazuju delimične adaptacije – recimo, veza između njih je prerasla u pun zglob koji omogućuje da se vrat fleksibilnije kreće u stranu. No, ograničen broj pršljenova ipak proizvodi čitav spektar problema, kao što vidimo na fotografiji žirafe koja pije: nezgrapna i ranjiva pozicija u kojoj se životinje nalaze kada god piju vodu ili pasu travu.
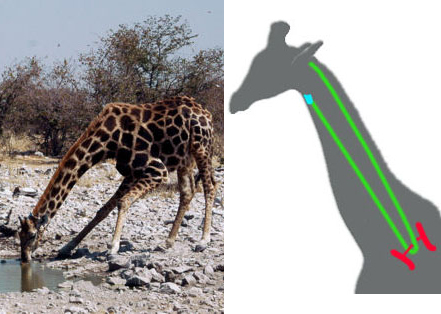
Levo: žirafin vrat sa samo sedam pršljenova predstavlja veliko ograničenje.
Desno: put rekurentnog laringijalnog nerva kroz vrat žirafe.
Zbog ovog ograničenja u broju vertebri, žirafe imaju ogromne probleme sa pokretljivošću vrata, pogotovo kada moraju da se sagnu da bi pile vodu. Starije žirafe često ginu na pojilu zato što ne mogu dovoljno brzo da se isprave kada budu napadnute od strane mesoždera. Ako se noge suviše rašire, žirafa pada na zemlju i više ne može da ustane.
Još jasniji zaostatak evolucionih procesa se vidi ako pogledamo jedan nerv u vratu, takozvani rekurentni laringijalni nerv, koji povezuje koren mozga sa delom grla poznatim kao larinks. Zbog razvojnog sistema koji je kroz evoluciju ostao očuvan još od riba (detaljan opis se može naći ovde), ovaj nerv u toku rasta prati iste signale koji upravljaju razvojem srčanih krvnih sudova. U vratu žirafe, ovo preuzima gigantske razmere: da bi povezao dve tačke udaljene samo nekoliko centimetara, rekurentni laringijalni nerv putuje više od šest metara.
Prsti konja
Tokom prvih pedeset godina prošlog veka, evolutivni niz u razvoju konja je bio među najbolje poznatima, tako da je često korišćen kao primer u udžbenicima biologije. Iz tog razloga se takođe često nalazio na meti antinaučnih napada, koji su napisali silne tekstove pokušavajući da negiraju postojanje fizičkih činjenica.
Tri tvrdnje su posebno česte. Prva je da su fosili Hyracotheriuma, prvog pretka konja, nađeni u istom sloju u kome su nađeni ostaci modernih konja. Da je ova tvrdnja istinita, ona ne bi u stvari bila nikakav problem po pitanju evolucije – kao što je objašnjeno u uvodu, originalna vrsta može da preživi i duže nego vrste koje su se odvojile od nje. No, ova tvrdnja je u svakom slučaju potpuno neistinita. Takvi nalazi prosto ne postoje. Ova neistina se pojavila u jednoj kreacionističkoj knjizi, i odatle je ljudi i dan danas kopiraju – bez ikakve provere, verujući autorima na reč.
Druga tvrdnja je da prelazni fosili konja zapravo nisu prelazni fosili pošto nema prelaznih fosila između prelaznih fosila (procenu logike koja stoji iza ovakvih stavova ostavljamo čitaocima). Recimo, jedan domaći kreacionista tvrdi da se “jednoprsti brstioci poput Equus-a javljaju iznenada, bez posrednika koji pokazuje postepenu evoluciju iz troprstih”.
Ovo je rezultat pisanja teksta o fosilima bez da se ti fosili prvo pogledaju. Shema A pokazuje razvoj zadnjeg kopita konja kroz nekoliko međustepena u stablu, slika B pokazuje kako se razvija kopito ždrebeta modernih konja. Jasno se vidi kako se dva prsta stapaju sa osnovnom kosti koja podržava jedini preostali prst (kopito).
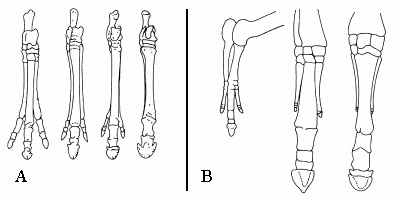
Treća tvrdnja je da su nađeni mnogi fosili koji se “ne uklapaju” u ovu shemu. Ova tvrdnja je tehnički sasvim tačna – standardna shema evolucije konja koja se koristi kao primer u udžbenicima biologije prikazuje direktnu evoluciju konja, bez drugih grana koje su se odvajale tokom te evolucije. No, kompletna slika daje još spektakularniji dokaz evolucije, pokazujući masu vrsta srodnih konjima koje su nekada postojale, i sve međukorake u njihovom razvoju. Za detaljnu sliku o istoriji evolucije konja, pogledajte ovde.
Kitovi: vodene krave, doslovno
Razvoj kitova počinje od mesožderskih kopitara iz izumrle grupe Pakicetidae. Ove životinje su bile grabljivci, koji su lovili plen u okolini reka ili u plitkoj vodi. Nisu bile dobri plivači, ali su bile prilagođene za lov u plićaku, uključujući i veoma specifičnu građu uha koja je omogućavala jasan sluh i na vodi i na suvom.
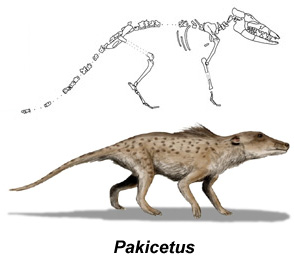
Autor rekonstrukcije: Nobu Tamura
Dalja adaptacija pakicetida na život u vodi se može videti kod sledećeg ključnog prelaznog fosila, Ambulocetus natans. Ovo stvorenje je imalo običan dugački sisarski rep, četiri kratke noge, zadnje noge sa velikim, proširenim stopalima sličnim perajima. Donekle postoji sličnost sa današnjim morskim lavovima, osim što je struktura skeleta očigledno bliska kitovima i delfinima. Lobanja veoma kitolika, sa tipičnom izduženom njuškom koju vidimo kod današnjih delfina. Nozdrve su još uvek na istom mestu kao i kod drugih sisara. Kroz analizu izotopa kiseonika, znamo da je Ambulocetus živeo i u morskoj i u slatkoj vodi, najverovatnije u ušćima velikih reka.
Veoma zanimljiv aspekt Ambulocetusa je njegov sluh: vide se prvi tragovi adaptacije vilične kosti za prenos zvuka. Ova povećana osetljivost na podvodne tonove je Ambulocetusu povećavala sposobnost lova, a dugoročno je postavila temelje za razvoj buduće eholokacije kod kitova i delfina.
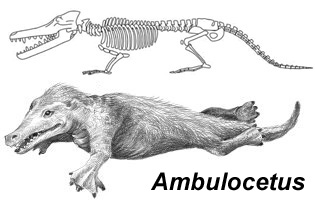
Skica fosila Pakicetusa: J.G.M. Thewissen. Autor rekonstrukcije: Nobu Tamura
Dalji razvoj možemo da pratimo u grupi vrsta poznatoj kao Remingtoncetidae. Iz fosilnog zapisa nam je poznato šest vrsta iz ove grupe, koje su cvetale u priobalnim područjima današnje južne Azije punih sedam miliona godina. Remingtoncetidae su postale potpuno morske životinje: za razliku od Ambulocetusa, njihovi udovi su postali kraći i zdepastiji, bolji za plivanje ali lošiji za hodanje po tlu. Takođe, u okviru njihove lobanje se dogodila još jedna ključna promena – polukružni kanali u unutrašnjem uhu, kritični organi za održavanje ravnoteže, su postali znatno manji. Mnogi paleontolozi smatraju da je ovo bila kritična tačka u evoluciji kitova: kada se ova promena jednom dogodila, dalja evolucija je mogla da teče samo u pravcu sve veće zavisnosti od mora.

Kutchicetus
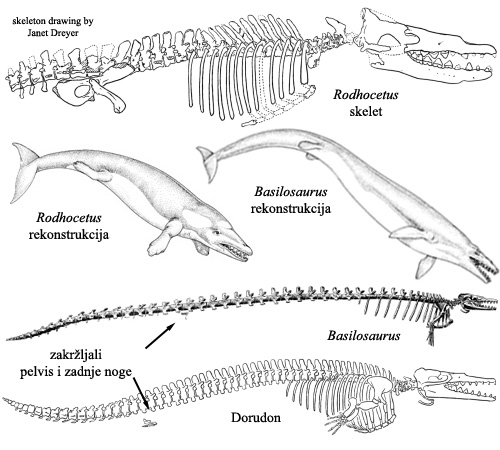
Sledeća grupa koja ilustruje nastanak kitova su Protocetidae. Najpoznatija vrsta iz ove grupe je Rodhocetus. Ovaj organizam je imao građu tela veoma sličnu Ambulocetusu, osim što su zadnje noge zakržljale skoro do neupotrebljivosti, a prednje postale nesposobne za hodanje po tlu. Rodhocetus je mogao da puzi po zemlji, ali nije više mogao da zaista hoda. Rep je postao masivan i adaptiran za plivanje.
Nakon ovog koraka, rani kitovi su se razgranali u raznim pravcima, sa raznim oblicima, veličinama, i dužinama. Među mnogim primerima iz ovog perioda je, na primer, Basilosaurus. Ova vrsta nije direktan predak današnjih kitova, ali je vrlo srodna, i njeni fosili pokazuju dalju specijalizaciju za život u dubokoj vodi. Radi se o skoro potpuno kitolikoj životinji, ako se ignoriše postojanje zakržljalih i sada već potpuno neupotrebljivih zadnjih udova. Rep pokazuje tragove postojanja repnog “peraja” kakvo imaju kitovi. Nozdrve se stapaju u jednu rupu koja se pomera više uz gornju vilicu.
Dorudontinae je grupa koja je živela u istom periodu kada i Basilosaurus, i verovatno jeste predak svih današnjih kitova. Poznato je devet različitih vrsta Dorudontinae, od kojih je na donjoj slici prikazana vrsta Dorudon atrax. Zadnje noge su još manje nego kod Basilosaurusa, druge osobine su skoro identične. Ista situacija je sa još nekoliko sličnih vrsta nađenih u ovom periodu: radi se o kitovima sa zadnjim nogama. Ove zadnje noge su, iako zakržljale, i dalje bile funkcionalne: koleno je još uvek postojalo i funkcionisalo, kao i prsti. Sluh se razvio još malo u poređenju sa Protocetidae, ali još uvek nije bilo eholokacije. Mozak je takođe i dalje bio veoma mali, uporediv sa mozgom njihovih predaka.
Od ove tačke sledi razvoj modernih Cetacea. Eholokacija se razvija kod prvih Odontocetes, kitova sa zubima, koji i dan danas koriste zube za proizvodnju eholokacionih signala; imamo fosile više desetina vrsta iz ovog prelaza. Nedugo nakon ovoga, razvija se grupa vrsta koje su paralelno imale i zube i takozvanu „kitovu kost“ (baleen) koju moderni kitovi koriste za filtriranje mikroskopskihi organizama iz morske vode. Primer ovoga je grupa Aetiocetus, u okviru koje imamo fosile četiri različite vrste. Vredi primetiti da na fosilima iz ovog perioda vidimo i pomeranje nozdrva uz lobanju, ka poziciji na vrhu glave koja postoji kod današnjih kitova.

Pomeranje nozdrva: sa leve strane, fosilna lobanja Pakicetusa, nozdrve su pri vrhu nosa. U sredini, fosilna lobanja Aetiocetusa, nozdrve su na sredini. Desno, lobanja modernog beluga kita, nozdrve su na vrhu lobanje.
Opet vredi napomenuti da su ovde u pitanju samo fosilni dokazi. Preci kitova i preci današnjih kopitara su potomci mesožderske grupe zvane mezonikidi. Genetski dokazi pokazuju razlaz upravo u periodu na koga ukazuju fosilni dokazi. Pored velikog broja fosilnih kitova sa zakržljalim nogama, i čak današnjih kitova koji još uvek imaju malu “ploveću” kost, poslednji ostatak pelvisa, tu su i drugi atavizmi: kitovi i dalje imaju ostatke sisarskog čula njuha, na primer.
Embriološki dokazi, paleoekološki dokazi, paleobiogeografski dokazi…sve informacije koje dobijamo iz ovih nezavisnih grana naučnog istraživanja potvrđuju zaključke izvedene na osnovu fosilnog zapisa.
Otkud glodari?
Evolucija glodara se u nekim izvorima prikazuje kao velika misterija, iako je to veoma daleko od istine. Osnovno grananje počinje od Euarchontoglires, grupe placentalnih sisara koji nisu bili daleko od stvorenja kao što je u prethodnom tekstu već pomenuta Eomaia. Ova početna grupa se ubrzo podelila na dve; jedna podgrupa Euarchontoglires, zvana Euarchonta, je postala predak današnjim redovima Scandentia (tupaja i slične male šumske insektivore) i primatima (među kojima su i ljudi). Druga podgrupa, zvana Glires, je postala predak današnjim lagomorfima (životinje kao što su zečevi) i glodarima.
Prve primere ovog prelaza nalazimo u kasnom Paleocenu i srednjem Eocenu sa grupom vrsta poznatom kao eurimiloidi. Danas imamo niz fosila pripadnika ove grupe kao što su Heomus, Sinomylus i Matutinia. Ova grupa pokazuje razvoj od korenskih insektivora: polako se gube očnjaci, zubi počinju da liče na tipične glodarske, gleđ postaje deblja sa prednje strane zuba, i javlja se razmak među prednjim zubima.
Sledi niz vrsta kao što je Tribosphenomys, kod koga vidimo konačni ishod razvoja glodarskih zuba (stalni rast prednjih zuba, i tipična građa). Acritoparamys je prvi primitivni glodar koji je potpuno poznat. Još uvek mnoge preostale osobine ranih sisara, ali imao noge više prilagođene za skakutanje.
Paramys i ostale vrste iskiromida (kasni Paleocen) su bili pra-glodari slični vevericama, ali bez adaptacije za život u drveću. Primitivna vilica (čije osobine ostaju i kod današnjih veverica), glodarski prednji zubi, ostali zubi još uvek slični primitivnim sisarima.
Od ove tačke počinje razlaz glodara na njihove osnovne podgrupe, i veoma bogat fosilni zapis prelaznih oblika.